氧循环
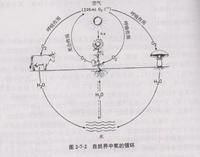
氧元素在自然界的循环状态。动植物的呼吸作用及人类活动中的燃烧都需要消耗氧气,产生二氧化碳。但植物的光合作用却大量吸收二氧化碳,释放氧气,如此构成了生物圈的氧循环(氧循环和碳循环是相互联系的).。氧在各圈层中的浓度如下(括号内数字为元素的丰度次序): 地球整体:28.5%(2) 地壳:46.6%(1)海洋:总量85.8%(1)溶解氧量,15℃时为6mg/kg(13) 大气:23.2%(2)。
-
选择特殊符号
选择搜索类型
请输入搜索
氧元素在自然界的循环状态。动植物的呼吸作用及人类活动中的燃烧都需要消耗氧气,产生二氧化碳。但植物的光合作用却大量吸收二氧化碳,释放氧气,如此构成了生物圈的氧循环(氧循环和碳循环是相互联系的).。氧在各圈层中的浓度如下(括号内数字为元素的丰度次序): 地球整体:28.5%(2) 地壳:46.6%(1)海洋:总量85.8%(1)溶解氧量,15℃时为6mg/kg(13) 大气:23.2%(2)。
一般在潜水泵的出水口的边上有一个插气管的头,将一根气管插在那上面另一头固定到水面以上,泵的出水口不插水管,开泵,你就会看到出水口有气泡喷出
在制冷空调技术中,目前最有希望取代制冷剂CFCiZ的是HFC一s4a,但是它的成本高,热物性尚不尽人意,润滑也不太理想。为了开拓其它替代物,文献〔1〕提出:采用跨临界制冷循环,以C02作制冷剂当是解决...
1.氧气泵也叫增氧泵、空气泵、打氧泵等,与工业增氧泵工作原理相似,一般功率较小。氧气泵的原理是将空气压入水中,让空气中的氧气与水体充分接触,使氧气融入水中,从而增加水体的溶氧量,以保证耗氧类生物的生长...

 臭氧发生器循环冷却水系统改造
臭氧发生器循环冷却水系统改造
臭氧发生器循环冷却水系统改造
臭氧发生器循环冷却水系统改造
 内循环厌氧反应器处理脂肪酸废水
内循环厌氧反应器处理脂肪酸废水
内循环厌氧反应器处理脂肪酸废水
研究了厌氧内循环反应器处理脂肪酸废水的启动和运行效果,化学需氧量(Chemical Oxygen De-mand,COD)与容积负荷的关系、容积负荷与出水挥发性脂肪酸(Volatile Fatty Acid,VFA)的关系.结果表明,反应器25 d即可完成启动,达到设计运行负荷25 kg COD/(m3.d);反应器运行负荷为25 kg COD/(m3.d)时,处理效果最佳,出水pH值为6.80~7.23,VFA约为130 mg/L,COD去除率达到85%以上;pH值变化滞后,VFA的变化比pH值能更好表征反应器内部的运行状况.
2015年10月9日,《臭氧处理循环冷却水技术规范》发布。
2016年5月1日,《臭氧处理循环冷却水技术规范》实施。
IC反应器(内循环厌氧反应器):
内循环(internalcirculation)厌氧反应器(简称IC反应器),是20世纪80年代中期荷兰PAQUES在UASB反应器(上流式厌氧污泥床)的基础上成功开发的第三代高效厌氧生物反应器。主要对水力流态进行较大改进而产生的新型反应器,该类型反应器不仅具有以往各代反应器生物量高的特点同时又具有良好的水力混合特性大大提高了反应器的处理能力,主要用于废水处理、污水治理、沼气生产等方面。
以上内容均根据学员实际工作中遇到的问题整理而成,供参考,如有问题请及时沟通、指正。
乙酰-CoA进入由一连串反应构成的循环体系,被氧化生成H₂O和CO₂。由于这个循环反应开始于乙酰CoA与草酰乙酸(oxaloaceticacid)缩合生成的含有三个羧基的柠檬酸,因此称之为三羧酸循环或柠檬酸循环(citratecycle)。在三羧酸循环中,柠檬酸合成酶催化的反应是关键步骤,草酰乙酸的供应有利于循环顺利进行。其详细过程如下:
1、乙酰-CoA进入三羧酸循环
乙酰CoA具有硫酯键,乙酰基有足够能量与草酰乙酸的羧基进行醛醇型缩合。首先柠檬酸合酶的组氨酸残基作为碱基与乙酰-CoA作用,使乙酰-CoA的甲基上失去一个H+,生成的碳阴离子对草酰乙酸的羰基碳进行亲核攻击,生成柠檬酰-CoA中间体,然后高能硫酯键水解放出游离的柠檬酸,使反应不可逆地向右进行。该反应由柠檬酸合酶(citratesynthase)催化,是很强的放能反应。由草酰乙酸和乙酰-CoA合成柠檬酸是三羧酸循环的重要调节点,柠檬酸合酶是一个变构酶,ATP是柠檬酸合酶的变构抑制剂,此外,α-酮戊二酸、NADH能变构抑制其活性,长链脂酰-CoA也可抑制它的活性,AMP可对抗ATP的抑制而起激活作用。
2、异柠檬酸形成
柠檬酸的叔醇基不易氧化,转变成异柠檬酸而使叔醇变成仲醇,就易于氧化,此反应由顺乌头酸酶催化,为一 可逆反应。
3、第一次氧化脱羧
在异柠檬酸脱氢酶作用下,异柠檬酸的仲醇氧化成羰基,生成草酰琥珀酸(oxalosuccinicacid)的中间产物,后者在同一酶表面,快速脱羧生成α-酮戊二酸(α-ketoglutarate)、NADH和CO2,此反应为β-氧化脱羧,此酶需要镁离子作为激活剂。此反应是不可逆的,是三羧酸循环中的限速步骤,ADP是异柠檬酸脱氢酶的激活剂,而ATP,NADH是此酶的抑制剂。
4、第二次氧化脱羧
在α-酮戊二酸脱氢酶系作用下,α-酮戊二酸氧化脱羧生成琥珀酰-CoA、NADH·H+和CO₂,反应过程完全类似于丙酮酸脱氢酶系催化的氧化脱羧,属于α-氧化脱羧,氧化产生的能量中一部分储存于琥珀酰coa的高能硫酯键中。α-酮戊二酸脱氢酶系也由三个酶(α-酮戊二酸脱羧酶、硫辛酸琥珀酰基转移酶、二氢硫辛酸脱氢酶)和五个辅酶(tpp、硫辛酸、hscoa、NAD+、FAD)组成。此反应也是不可逆的。α-酮戊二酸脱氢酶复合体受ATP、GTP、NADH和琥珀酰-CoA抑制,但其不受磷酸化/去磷酸化的调控。
5、底物磷酸化生成ATP
在琥珀酸硫激酶(succinatethiokinase)的作用下,琥珀酰-CoA的硫酯键水解,释放的自由能用于合成gtp,在细菌 和高等生物可直接生成ATP,在哺乳动物中,先生成GTP,再生成ATP,此时,琥珀酰-CoA生成琥珀酸和辅酶A。
6、琥珀酸脱氢
琥珀酸脱氢酶(succinatedehydrogenase)催化琥珀酸氧化成为延胡索酸。该酶结合在线粒体内膜上,而其他三羧酸循环的酶则都是存在线粒体基质中的,这酶含有铁硫中心和共价结合的FAD,来自琥珀酸的电子通过FAD和铁硫中心,然后进入电子传递链到O₂,丙二酸是琥珀酸的类似物,是琥珀酸脱氢酶强有力的竞争性抑制物,所以可以阻断三羧酸循环。
7、延胡索酸的水化
延胡索酸酶仅对延胡索酸的反式双键起作用,而对顺丁烯二酸(马来酸)则无催化作用,因而是高度立体特异性的。
8、草酰乙酸再生
在苹果酸脱氢酶(malicdehydrogenase)作用下,苹果酸仲醇基脱氢氧化成羰基,生成草酰乙酸(oxalocetate),NAD+是脱氢酶的辅酶,接受氢成为NADH·H+(图4-5)。
在此循环中,最初草酰乙酸因参加反应而消耗,但经过循环又重新生成。所以每循环一次,净结果为1个乙酰基通过两次脱羧而被消耗。循环中有机酸脱羧产生的二氧化碳,是机体中二氧化碳的主要来源。在三羧酸循环中,共有4次脱氢反应,脱下的氢原子以NADH+H+和FADH2的形 式进入呼吸链,最后传递给氧生成水,在此过程中释放的能量可以合成ATP。乙酰辅酶A不仅来自糖的分解,也可由脂肪酸和氨基酸的分解代谢中产生,都进入三羧酸循环彻底氧化。并且,凡是能转变成三羧酸循环中任何一种中间代谢物的物质都能通过三羧酸循环而被氧化。所以三羧酸循环实际是糖、脂、蛋白质等有机物在生物体内末端氧化的共同途径。三羧酸循环既是分解代谢途径,但又为一些物质的生物合成提供了前体分子。如草酰乙酸是合成天冬氨酸的前体,α-酮戊二酸是合成谷氨酸的前体。一些氨基酸还可通过此途径转化成糖。


