核酸、蛋白质谁更"牛"?
一般人都知道,生命是蛋白质存在的形式,蛋白质是生命的基础。在发现核酸前,这句话是对的,但当核酸被发现后,应该说最本质的生命物质是核酸,或是把上述的这句话更正为蛋白体是生命的基础。按照现代生物学的观点,蛋白体是包括核酸和蛋白质的生物大分子。
核酸在生命中为什么比蛋白质更重要呢?因为生命的重要性是能自我复制,而核酸就能够自我复制。蛋白质的复制是根据核酸所发出的指令,使氨基酸根据其指定的种类进行合成,然后再按指定的顺序排列成所需要复制的蛋白质。世界上各种有生命的物质都含有蛋白体,蛋 白体中有核酸和蛋白质,至今还没有发现有蛋白质而没有核酸的生命。但在有生命的病毒研究中,却发现病毒以核酸为主体,蛋白质和脂肪以及脂蛋白等只不过充作其外壳,作为与外界环境的界限而已,当它钻入寄生细胞繁殖子代时,把外壳留在细胞外,只有核酸进入细胞内 ,并使细胞在核酸控制下为其合成子代的病毒。这种现象,美国科学家比喻为人和汽车的关系。即把核酸比为人,蛋白质比作汽车,人驾驶汽车到处跑,外表上看,人车一体是有生命运动的东西,而真正的生命是人,汽车只是由人制造的载入的外壳。有一种类病毒,是能繁殖子代的有生命物体,其中只有核酸而没蛋白质,可见核酸是真正的生命物质。
因此中国1996年最新出版的《人体生理学》改变了旧教科书中只提蛋白质是生命基础的缺陷,明确提出:"蛋白质和核酸是一切生命活动的物质基础。"
然而,多少年来,人们在一味追求蛋白质、维生素、微量元素等营养时,却把最重要的角色--核酸忘却了,这不能不说是人类生命史上的一大遗憾。
没有核酸,就没有蛋白,也就没有生命。
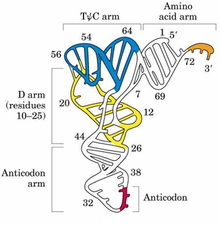
核酸也称多聚核苷酸,是由许多个核苷酸聚合而成的生物大分子,核苷酸是由含氮的碱基、核糖或脱氧核糖、磷酸三种分子连接而成。碱基与糖通过糖苷键连接成核苷,核苷与磷酸以酯键连接成核苷酸。核苷酸是生物体内一类重要含氮化合物,是各种核酸的基本组成单位。根据核酸所含戊糖的不同,可分为核糖核酸(RNA)和脱氧核糖核酸(DNA)二种。
核酸不但是一切生物细胞的基本成分,还对生物体的生长、发育、繁殖、遗传及变异等重大生命现象起主宰作用。它在生物科学的地位,可用"没有核酸就没有生命"这句话来概括。
饮食核酸的营养保健作用如下:
1.饮食核酸与免疫 '
从核酸对机体各系统的影响来看,免疫系统是最敏感也是最直接受影响的系统。1985年科学家就证实无核酸饮食或低核酸饮食配方饲喂的实验动物,其细胞免疫功能低下,条件致病菌就可使其感染。无核酸饮食致使T淋巴细胞发育障碍、功能低下,而没有细胞免疫反应的发生,同时影响T细胞依赖的体液免疫的产生;补充核酸营养后可恢复免疫系统的发育和免疫功能。实验表明,核酸是维持机体正常免疫功能和免疫系统生长代谢的必需营养物质。
2.饮食核酸与衰老和内分泌
衰老是机体各组织器官的退行性变化,关于衰老发生机制的学说很多,如自由基学说、免疫学说、内分泌学说、遗传学说等。脂质过氧化随年龄增大而增高,并伴有酶与非酶系统防御功能下降,导致体内自由基浓度升高。代谢性、退行性疾病的发生和发展与体内过氧化脂质含量高度正相关。饮食核酸能增加血浆单不饱和脂肪酸和co-3、¨6系列多不饱和脂肪酸的含量,多不饱和脂肪酸的增加可提高机体对抗自由基的能力。饮食核酸作为使遗传物质活泼代谢的原料,具有极强的抗生物氧化、消除体内自由基和全面增强免疫功能及性激素分泌的作用,因此在延缓衰老方面优势显著。
3.饮食核酸与增殖细胞
饮食中补加核酸有助于肝脏再生和受损伤的小肠恢复功能。有无核酸饮食对比研究证明,一段时期内膳食中如缺乏核酸,将对大鼠肝脏的超微结构及功能造成不良影响,提示饮食核酸是维持肝脏处于正常生理状态的必需营养物质。血液中的红细胞、白细胞、血小板和血浆蛋白等也都是代谢较快的增殖细胞系,加之它们中的大多数均无从头合成核酸的能力,因此它们的代谢和功能也都需要充足的核酸营养。再生障碍性贫血和抗癌药物、放疗、化疗等引起的贫血,即缺铁性贫血之外的贫血均需补充核酸营养,以改善骨髓造血功能和血液成分的代谢活力。
4.饮食核酸与癌症
人体每日约有数百万个癌状细胞出现,它们几乎全部被机体的免疫监视系统和核酸、维生素等食物成分,在形成大的癌细胞克隆前排除掉。因此在日常生活中尽量避免致癌因子的作用,增加核酸等防癌因素的作用非常必要。
5.饮食核酸与痴呆等神经障碍
食物核酸提取物对痴呆症状的改善非常令人鼓舞。在大鼠实验中,如给大鼠脑注射RNA合成阻断剂,则所掌握的学习能力和记忆能力在5小时后丧失,但如在注射RNA合成阻断剂的同时注射拮抗阻断剂的物质,这种记忆丧失就不发生。美国哈佛大学的研究也表明,老年痴呆患者脑内神经纤维变化多的部位,RNA和蛋白质合成显著减少,因此发生记忆障碍。
6.饮食核酸与循环系统
核酸营养对循环系统的作用是抑制过氧化脂质的形成,抑制胆固醇的生成,扩张血管,改善血流,纠正心肌代偿不良,促进血管壁再生,抑制血小板凝集i因此核酸被认为对脑血栓、心肌梗死、高血压和动脉粥样硬化症有较好的营养保健作用。
7、饮食核酸与糖尿病
非胰岛素依赖性糖尿病与生活方式和运动不足关系密切,如果在普通的饮食疗法的基础上,再加上核酸饮食,将收到更好的效果。其原因:一是糖尿病患者血清中过氧化脂质增多,核酸及其代谢产物对其具有较强的清除作用;二是由于核酸的促细胞(包括促胰脏的胰岛素分泌细胞)代谢功能。除此之外,核酸的代谢产物腺苷还有抑制糖的分解作用,使糖在小肠内的吸收减缓。
除上述作用外,饮食核酸还有以下作用:减肥,提高机体对环境变化的耐受力,显著的抗疲劳、增强机体对冷热的抵抗力、促进摄人氧气的利用,促进小鼠生殖系统的发育等。
对于婴儿、迅速成长期的孩子、老年体弱多病、全身感染、外伤手术者、肝功能不全以及白细胞、T细胞、淋巴细胞降低人群等,可以额外补充核酸类物质。世界卫生组织规定,每天膳食中核酸的量不大于2克,扣除食物中的核酸摄入量,每天补充小于1.5克核酸是合适的。
人工核酸治疗白血病
日本工业技术院产业技术融合领域研究所已开发出了治疗白血病的人造核酸。这种人造核酸就像一把剪刀,可发现引起白血病的遗传基因并将其剪除。科研小组的成员、东京大学研究生院教授多比良和诚根据动物实验结果认为,这种人造核酸将来有望成为治疗白血病的主要药物。
这次研究的对象是慢性骨髓性白血病(MCL),患者的异常遗传因子是由两个正常的遗传因子连接而成的,新开发的人造核酸可以发现这种变异遗传基因并将其切断。科学家过去也发现过能找到特定的遗传因子序列并将其切断的分子,但在切断特定遗传因子序列的同时往往对正常细胞造成伤害。而新开发出的核酸只在发现异常遗传因子时才被激活,平时则潜伏不动。
科研小组用人体白血病细胞进行了动物实验。他们将可与人造核酸反应的细胞和不可与人造核酸反应的细胞分别注射到8只实验鼠的体内。移植后第13周时,不与人造核酸反应的细胞全部死亡,而与人造核酸反应的细胞全部存活,证明人造核酸在生物体内十分有效。
科研小组说,此人造核酸的临床应用尚有诸多问题要解决,将来很可能是把患者的骨髓细胞抽出来,经人造核酸处理后,再把正常细胞的骨髓输回患者体内。
核酸的生物功能
核酸在生物体内主要与蛋白质合成核蛋白存在,它既是蛋白质生物合成不可缺少的物质,又是生物遗传的物质基础。
DNA主要存在于细胞核中,它们是遗传信息的携带者,DNA的结构决定生物合成蛋白质的特定结构,并保证这种遗传特性传给下一代。RNA主要存在于细胞质中,它们是以DNA为模板而形成的,并且直接参加蛋白质的生物合成过程。因此,DNA是RNA的模板,而RNA又是蛋白质的模板。存在于DNA分子上的遗传信息就是这样由DNA传递给RNA,再传递给蛋白质。通过DNA 复制,遗传信息一代代传下去,正因为有这样的功能,人们把核酸誉为"生命之源"和"生命之本"。