红树林和互花米草相互作用对环境变化的多尺度响应项目摘要文献

 红树林湿地公园项目建设汇报材料
红树林湿地公园项目建设汇报材料
红树林湿地公园项目建设汇报材料
红树林湿地公园项目建设汇报材料 按照区委办下发的《关于重新印发 <海陵区重大项目工 作组一览表 >》的文件通知,神前湾红树林湿地公园项目于 2013年 11 月 15日被列为区重点项目。根据 2013年 10月 31 日海陵岛试验区下发的 《区管委主任办公会议纪要》 第三 条“听取红树林湿地公园项目规划情况汇报”的指示,由海 陵区旅游文体局为该项目业主单位,统一负责该项目的建设 和管理。 目前,全区红树林面积覆盖率最大片区为神前湾浅海滩 涂红树林,现有红树林 220亩,近期补种 150亩。其特性是 消浪护堤,促淤保滩。神前湾红树林北靠灵猴山,南依兰章 联围,东至神前村,西临灵谷港、灵谷庙。神前湾红树林片 区地理位臵优越,无论在生态保护还是旅游开发,都是一个 天然的合适地点。为进一步增强我区的原生态旅游功能,营 造舒适、美观、安全、原生态的旅游氛围,丰富旅游项目和 营造良好投资环境,从而提升
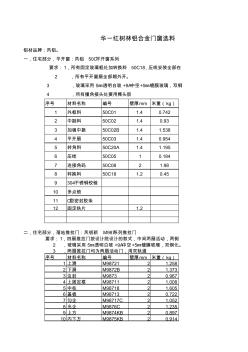 华一红树林门窗选料
华一红树林门窗选料
华一红树林门窗选料
序号 材料名称 编号 壁厚mm 米重( kg) 1 外框料 50C01 1.4 0.742 2 中挺料 50C02 1.4 0.93 3 加强中挺 50C02B 1.4 1.538 4 平开扇 50C03 1.4 0.954 5 转角料 50C20A 1.4 1.195 6 压线 50C05 1 0.184 7 连接角码 50C08 2 1.98 8 转换料 50C18 1.2 0.45 9 304不锈钢铰链 10 多点锁 11 O型密封胶条 12 固定铁片 1.2 序号 材料名称 编号 壁厚mm 米重( kg) 1 上滑 M98721 2 1.258 2 下滑 M9872B 2 1.373 3 边封 M9873 2 0.967 4 上固定框 M98711 2 1.008 5 中柱 M98718 2 1.605 6 盖板 M98713 2 0.722 7 勾企 M98717C 2 1.05