白及属
Bletilla Rchb. f.
地生植物。茎基部具膨大的假鳞茎,其近旁常具多枚前一年和以前每年所残留的扁球形或扁卵圆形的假鳞茎;假鳞茎的侧边常具2枚突起,彼此以同一方向的突起与毗邻的假鳞茎相连成一串,假鳞茎上具荸荠似的环带,肉质,富粘性,生数条细长根。叶(2-)3-6枚,互生,狭长圆状披针形至线状披针形,叶片与叶柄之间具关节,叶柄互相卷抱成茎状。花序顶生,总状,常具数朵花,通常不分枝或极罕分枝;花序轴常常曲折成“之”字状;花苞在开花时常凋落;花紫红色、粉红色、黄色或白色,倒置,唇瓣位于下方;萼片与花瓣相似,近等长,离生;唇瓣中部以上常明显3裂;侧裂片直立,多少抱蕊柱,唇盘上从基部至近先端具5条纵脊状褶片,基部无距;蕊柱细长,无蕊柱足,两侧具翅,顶端药床的侧裂片常常为略宽的圆形,后侧的中裂片齿状;花药着生于药床的齿状中裂片上,帽状,内屈或者近于悬垂,具或多或少分离的2室;花粉团8个,成2群,每室4个,成对而生,粒粉质,多颗粒状,具不明显的花粉团柄,无粘盘;柱头1个,位于蕊喙之下。蒴果长圆状纺锤形,直立。
约6种,分布于亚洲的缅甸北部经中国至日本。中国产4种,分布于北起江苏、河南,南至台湾,东起浙江,西至西藏东南部(察隅)。
本属模式种: Bletilla gebina (Lindl.) Rchb. f. [Bletilla striata (Thunb. ex A. Murray) Rchb. f.]
石豆兰属
Bulbophyllum Thou.
附生草本。根状茎匍匐,少有直立的,具或不具假鳞茎。假鳞茎紧靠,聚生或疏离,形状、大小变化甚大,具1个节间。叶通常1枚,少有2-3枚,顶生于假鳞茎,无假鳞茎的直接从根状茎上发出;叶片肉质或革质,先端稍凹或锐尖、圆钝,基部无柄或具柄。花葶侧生于假鳞茎基部或从根状茎的节上抽出,比叶长或短,具单花或多朵至许多花组成为总状或近伞状花序;花苞片通常小;花小至中等大;萼片近相等或侧萼片远比中萼片长,全缘或边缘具齿、毛或其他附属物;侧萼片离生或下侧边缘彼此粘合,或由于其基部扭转而使上下侧边缘彼此有不同程度的粘合或靠合,基部贴生于蕊柱足两侧而形成囊状的萼囊;花瓣比萼片小,全缘或边缘具齿、毛等附属物;唇瓣肉质,比花瓣小,向外下弯,基部与蕊柱足末端连接而形成活动或不动的关节;蕊柱短,具翅,基部延伸为足;蕊柱翅在蕊柱中部或基部以不同程度向前扩展,向上伸延为形状多样的蕊柱齿;花药俯倾,2室或由于隔膜消失而成1室;花粉团蜡质,4个成2对,无附属物。
约1000种,分布于亚洲、美洲、非洲等热带和亚热带地区,大洋洲也有。中国有98种和3变种,主要产于长江流域及其以南各省区。
本属模式种:Bulbophyllum nutans Thou.
虾脊兰属
Calanthe R. Br.
地生草本。根圆柱形,细长而密被淡灰色长绒毛。根状茎有或无。假鳞茎通常粗短,圆锥状,很少不明显或伸长为圆柱形的。叶少数,常较大,少有狭窄而呈剑形或带状,幼时席卷,全缘或波状,基部收窄为长柄或近无柄,柄下为鞘,在叶柄与鞘相连接处有一个关节或无,无毛或有毛,花期通常尚未全部展开或少有全部展开的。花葶出自当年生由低出叶和叶鞘所形成的假茎上端的叶丛中,或侧生于茎的基部,少有从去年生无叶的茎端发出,直立,不分枝,下部具鞘或鳞片状苞片,通常密被毛,少数无毛;总状花序具少数至多数花;花苞片小或大,宿存或早落;花通常张开,小至中等大;萼片近相似,离生;花瓣比萼片小;唇瓣常比萼片大而短,基部与部分或全部蕊柱翅合生而形成长度不等的管,少有贴生在蕊柱足末端而与蕊柱分离的,分裂或不裂,有距或无距;唇盘具附属物(胼胝体、褶片或脊突)或无附属物;蕊柱通常粗短,无足或少数具短足,两侧具翅,翅向唇瓣基部延伸或不延伸;蕊喙分裂或不分裂;柱头侧生;花粉团蜡质,8个,每4个为一群,近相等或不相等;花粉团柄明显或不明显,共同附着于1个粘质物上。
全属约150种,分布于亚洲热带和亚热带地区、新几内亚岛、澳大利亚、热带非洲以及中美洲。中国有49种及5变种,主要产长江流域及其以南各省区。
本属模式种:Calanthe veratrifolia R. Br.(Orchis triplicata Willem., Calanthe triplicata (Willem.) Ames)
隔距兰属
Cleisostoma Bl.
附生草本。茎长或短,质地硬,直立或下垂,少有匍匐,分枝或不分枝,具多节。叶少数至多数,质地厚,二列,扁平,半圆柱形或细圆柱形,先端锐尖或钝并且不等侧2裂,基部具关节和抱茎的叶鞘。总状花序或圆锥花序侧生,具多数花;花苞片小,远比花梗和子房短;花小,多少肉质,开放,萼片离生,侧萼片常歪斜,花瓣通常比萼片小;唇瓣贴生于蕊柱基部或蕊柱足上,基部具囊状的距,3裂,唇盘通常具纵褶片或脊突;距内具纵隔膜,在内面背壁上方具1枚形状多样的胼胝体;蕊柱粗短,常金字塔状,具短的蕊柱足或无;蕊喙小;药帽前端伸长或不伸长;花粉团蜡质,4个,每不等大的2个为一对,具形状多样的粘盘柄和粘盘。
约100种,分布于热带亚洲至大洋洲。中国有17种和1变种,主要产于南方诸省区。
本属模式种:Cleisostoma sagittatum Bl.
贝母兰属
Coelogyne Lindl.
附生草本。根状茎常延长,匍匐或多少悬垂,通常具较密生的节,节上被鳞片状鞘。假鳞茎常较粗厚,以一定距离着生于根状茎上,基部常被箨状鞘,顶端生 (1-) 2枚叶。叶通常长圆形至椭圆状披针形,一般质地较厚,基部具柄。花葶生于假鳞茎顶端,常与幼叶同时出现于幼嫩假鳞茎顶端,初时发自近老假鳞茎基部的根状茎上,有时在花葶基部或上部近花序轴处具多枚二列套叠的、宿存的革质颖片;总状花序直立或俯垂,通常具数朵花,较少超过20朵或减退为单花的;花苞片常较大,近舟状,对折,脱落;花较大或中等大,常白色或绿黄色,唇瓣多有斑纹;萼片相似,有时背面有龙骨状突起;花瓣常为线形,较少与萼片近等宽;唇瓣常以狭窄的基部着生于蕊柱基部,3裂或罕有不裂;侧裂片直立并多少围抱蕊柱;唇盘上有2-5条纵褶片或脊,后者常分裂或具附属物;蕊柱较长,上端两侧常具翅,翅可围绕蕊柱顶端,无蕊柱足;花药内倾;花粉团4个,成2对,蜡质,附着于1个粘质物上;柱头凹陷;蕊喙较大。蒴果中等大,常有棱或狭翅。
全属约200种,分布于亚洲热带和亚热带南缘至大洋洲。中国有26种,主要产西南、少数也见于华南。
本属后选模式种:Coelogyne cristata Lindl.
兰属
Cymbidium Sw.
附生或地生草本,罕有腐生,通常具假鳞茎;假鳞茎卵球形、椭圆形或梭形,较少不存在或延长成茎状,通常包藏于叶基部的鞘之内。叶数枚至多枚,通常生于假鳞茎基部或下部节上,二列,带状或罕有倒披针形至狭椭圆形,基部一般有宽阔的鞘并围抱假鳞茎,有关节。花葶侧生或发自假鳞茎基部,直立、外弯或下垂;总状花序具数花或多花,较少减退为单花;花苞片长或短,在花期不落;花较大或中等大;萼片与花瓣离生,多少相似;唇瓣3裂,基部有时与蕊柱合生达3-6毫米;侧裂片直立,常多少围抱蕊柱,中裂片一般外弯;唇盘上有2条纵褶片,通常从基部延伸到中裂片基部,有时末端膨大或中部断开,较少合而为一;蕊柱较长,常多少向前弯曲,两侧有翅,腹面凹陷或有时具短毛,花粉团2个,有深裂隙,或4个而形成不等大的2对,蜡质,以很短的、弹性的花粉团柄连接于近三角形的粘盘上。
全属约48种,分布于亚洲热带与亚热带地区,向南到达新几内亚岛和澳大利亚。中国有29种,广泛分布于秦岭山脉以南地区。
本属后选模式种:Cymbidium aloifolium (L.) Sw. (Epidendrum aloifolium L.)
杓兰属
Cypripedium L.
地生草本,具短或长的横走根状茎和许多较粗厚的纤维根。茎直立,长或短,成簇生长或疏离,无毛或具毛,基部常有数枚鞘。叶2至数枚,互生、近对生或对生,有时近铺地;叶片通常椭圆形至卵形,较少心形或扇形,具折扇状脉、放射状脉或3-5条主脉,有时有黑紫色斑点。花序顶生,通常具单花或少数具2-3花,极罕具5-7花;花苞片通常叶状,明显小于叶,少有非叶状或不存在;花大,通常较美丽;中萼片直立或俯倾于唇瓣之上;2枚侧萼片通常合生而成合萼片,仅先端分离,位于唇瓣下方,极罕完全离生;花瓣平展、下垂或围抱唇瓣,有时扭转;唇瓣为深囊状,球形、椭圆形或其他形状,一般有宽阔的囊口,囊口有内弯的侧裂片和前部边缘,囊内常有毛;蕊柱短,圆柱形,常下弯,具2枚侧生的能育雄蕊、1枚位于上方的退化雄蕊和1个位于下方的柱头;花药2室,具很短的花丝;花粉粉质或带粘性,但不粘合成花粉团块;退化雄蕊通常扁平,椭圆形、卵形或其他形状,有柄或无柄,极罕舌状或线形;柱头肥厚,略有不明显的3裂,表面有乳突。果实为蒴果。
本属后选模式种:Cypripedium calceolus L.
全属约50种,主要产东亚、北美、欧洲等温带地区和亚热带山地,向南可达喜马拉雅地区和中美洲的危地马拉。中国有32种,广布于自东北地区至西南山地和台湾高山,绝大多数种类均可供观赏。
石斛属
Dendrobium Sw.
附生草本。茎丛生,少有疏生在匍匐茎上的,直立或下垂,圆柱形或扁三棱形,不分枝或少数分枝,具少数或多数节,有时1至数个节间膨大成种种形状,肉质(亦称假鳞茎)或质地较硬,具少数至多数叶。叶互生,扁平,圆柱状或两侧压扁,先端不裂或2浅裂,基部有关节和通常具抱茎的鞘。总状花序或有时伞形花序,直立,斜出或下垂,生于茎的中部以上节上,具少数至多数花,少有退化为单朵花的;花小至大,通常开展;萼片近相似,离生;侧萼片宽阔的基部着生在蕊柱足上,与唇瓣基部共同形成萼囊;花瓣比萼片狭或宽;唇瓣着生于蕊柱足末端,3裂或不裂,基部收狭为短爪或无爪,有时具距;蕊柱粗短,顶端两侧各具1枚蕊柱齿,基部具蕊柱足;蕊喙很小;花粉团蜡质,卵形或长圆形,4个,离生,每2个为一对,几无附属物。
约1000种,广泛分布于亚洲热带和亚热带地区至大洋洲。中国有74种和2变种,产秦岭以南诸省区,尤其云南南部为多。
本属模式种:Dendrobium moniliforme (L.). Sw. (Epidendrum moniliforme L.)
羊耳蒜属
Liparis L. C. Rich.
地生或附生草本,通常具假鳞茎或有时具多节的肉质茎。假鳞茎密集或疏离,外面常被有膜质鞘。叶1至数枚,基生或茎生(地生种类),或生于假鳞茎顶端或近顶端的节上(附生种类),草质、纸质至厚纸质,多脉,基部多少具柄,具或不具关节。花葶顶生,直立、外弯或下垂,常稍呈扁圆柱形并在两侧具狭翅;总状花序疏生或密生多花;花苞片小,宿存;花小或中等大,扭转;萼片相似,离生或极少两枚侧萼片合生,平展,反折或外卷;花瓣通常比萼片狭,线形至丝状;唇瓣不裂或偶见3裂,有时在中部或下部缢缩,上部或上端常反折,基部或中部常有胼胝体,无距;蕊柱一般较长,多少向前弓曲,罕有短而近直立的,上部两侧常多少具翅,极少具4翅或无翅,无蕊柱足;花药俯倾,极少直立;花粉团4个,成2对,蜡质,无明显的花粉团柄和粘盘。蒴果球形至其他形状,常多少具3钝棱。
本属模式种: Liparis loeselii (L.) L. C. Rich. (Ophrys loeselii L.)
全属约有250种,广泛分布于全球热带与亚热带地区,少数种类也见于北温带。中国有52种。
毛兰属
Eria Lindl.
附生植物,通常具根状茎。茎常膨大成种种形状的假鳞茎,少有不膨大者,具1至多节,基部被鞘。叶1至数枚,通常生于假鳞茎顶端或近顶端的节上,较少在不膨大的茎上呈二列排列或散生于茎上。花序侧生或顶生,常排列成总状,较少减退为单花,被绵毛或无毛;花苞片小或稍大;花通常较小,少有较大并具鲜艳色彩;萼片背面与子房被绒毛或无毛;萼片离生;侧萼片多少与蕊柱足合生成萼囊;花瓣与中萼片相似或较小;唇瓣生于蕊柱足末端,具或不具关节,无距,常3裂,上面通常有纵脊或胼胝体;蕊柱短或长,具长短不同的蕊柱足;花药为不完全的4室;花粉团8个,每4个成一群,蜡质,基部收狭成柄状,附着于粘盘上。蒴果圆柱形。
全属约370余种,分布于亚洲热带至大洋洲。中国有43种,产南部各省区。
本属模式种:Eria stellata Lindl.
兜被兰属
Neottianthe Schltr.
地生草本。块茎圆球形或椭圆形,肉质,不裂,颈部生几条细长根。叶1或2枚,基生或茎生。花序顶生,总状,常具几条至多朵花,罕仅1(-2)朵花;花苞片直立伸展;花通常小,罕大,紫红色、粉红色或近白色,罕淡黄色或黄绿色,常偏向一侧,倒置(唇瓣位于下方);萼片近等大,彼此在3/4以上紧密靠合成兜;花瓣线形、线状披针形或长圆形,常较萼片稍短而狭,与中萼片贴生,罕与萼片近等宽;唇瓣向前伸展,从基部向下反折,常3裂,极罕在一株植物花序的某些朵花中唇瓣的侧裂片再2裂,唇瓣成4裂或5裂,上面具极密的细乳突,中裂片线形、线状舌形、长方形、披针形或卵形,侧裂片常较中裂片短而窄,基部具距;蕊柱短,直立;花药直立,长圆形或椭圆形,2室,先端钝,药室并行;花粉团2个,为具小团块的粒粉质,具短的花粉团柄和粘盘,粘盘小,卵形、近圆形或椭圆形,裸露;蕊喙小,隆起,三角形,位于药室基部之间;柱头2个,隆起,多少呈棍棒状,位于蕊喙之下;退化雄蕊2个,较小,近圆形,位于花药基部两侧。蒴果直立,无喙。
本属约12种,主要分布于亚洲亚热带至北温带山地,个别种分布至欧洲。中国产12种,四川和云南是其现代的分布中心和分化中心。
本属模式种:Neottianthe cucullata (L.) Schltr. (Orchis cucullata L. )
红门兰属
Orchis L.
地生草本,基部具细、指状、肉质的根状茎或具1-2枚肉质,长圆形、椭圆形、卵形或圆球形的块茎,块茎不裂或下部呈掌状分裂,裂片细长,颈部常有几条细长而多少弯曲的根。茎直立,圆柱形,靠近基部具1-3枚筒状鞘,鞘之上具叶,叶基生或茎生,互生,罕近对生,1-5枚,稍肥厚,基部收狭成鞘抱茎。总状花序顶生,具1至多数花,花偏向一侧或不偏向一侧,花序轴无毛或被短柔毛;花苞片常直立伸展,披针形、卵状披针形、卵形或倒披针形,短于或长于花;子房扭转,无毛或被短柔毛,具短或较长的花梗;花较小或中等大,少数较大,粉红色、紫红色、白色、黄绿色或黄色,花倒置(唇瓣位于下方);萼片离生,近等长或侧萼片较中萼片稍长,中萼片直立,常凹陷呈舟状,侧萼片张开或反折;花瓣与中萼片等长或较短小,常与中萼片相靠合呈兜状,边缘无睫毛或具睫毛;唇瓣常向前伸展,绝大多数种与花瓣的形状明显不一样,少数种则一样,不裂或3裂至4裂,边缘无睫毛或具睫毛,基部无爪或收狭具爪,基部有距,罕无距;距圆筒状或囊状;蕊柱直立,花药位于蕊柱顶部,药隔稍宽或较窄,顶部钝或稍突出,药室2,并行;花粉团2个,为具小团块的粒粉质,具花粉团柄和粘盘;粘盘2个,各埋藏于1个粘质球内,两个粘质球一起被包藏于蕊喙的1个粘囊中,粘囊通常为近球形,突出于蕊柱前面距口之上方;柱头1个,凹陷,位于蕊喙之下的凹穴内;蕊喙位于花药下部两药室之间;退化雄蕊2枚,位于花药基部两侧。蒴果直立。
本属后选模式种:Orchis militaris L.
约80种(广义),分布于北温带、亚洲亚热带山地以及北非的温暖地区。中国有28种,2变种,除南部炎热地区外,南北均产,尤以西南部高海拔山地为多。
天麻属
Gastrodia R. Br.
腐生草本,地下具根状茎;根状茎块茎状、圆柱状或有时多少呈珊瑚状,通常平卧,稍肉质,具节,节常较密。茎直立,常为黄褐色,无绿叶,一般在花后延长,中部以下具数节,节上被筒状或鳞片状鞘。总状花序顶生,具数花至多花,较少减退为单花;花近壶形、钟状或宽圆筒状,不扭转或扭转;萼片与花瓣合生成筒,仅上端分离;花被筒基部有时膨大成囊状,偶见两枚侧萼片之间开裂;唇瓣贴生于蕊柱足末端,通常较小,藏于花被筒内,不裂或3裂;蕊柱长,具狭翅,基部有短的蕊柱足;花药较大,近顶生;花粉团2个,粒粉质,通常由可分的小团块组成,无花粉团柄和粘盘。
全属约20种,分布于东亚、东南亚至大洋洲。中国有13种。
本属模式种:Gastrodia sesamoides R. Br.

 Hitachi的UAX电梯介绍
Hitachi的UAX电梯介绍
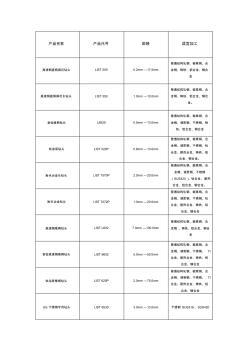 NACHI钻头参数
NACHI钻头参数