生物乙醇
生物乙醇是指通过微生物的发酵将各种生物质转化为燃料酒精。它可以单独或与汽油混配制成乙醇汽油作为汽车燃料。
-

选择特殊符号
选择搜索类型
请输入搜索
生物乙醇是指通过微生物的发酵将各种生物质转化为燃料酒精。它可以单独或与汽油混配制成乙醇汽油作为汽车燃料。
醇类按说没有酸性,是中性物质。但是如果羟基所连碳原子上有吸引电子的取代基,会使羟基上的氢原子电离程度增大;如果羟基所连碳原子上有排斥电子的取代基,会使羟基上的氢原子电离程度减小。氯原子是吸引电子的取代...
制备方法制备原料有淀粉、乙烯、磷酸、、葡糖淀粉酶,衍生产品为盐酸乙醇液、二硫化硒、环氧乙烷、对二乙基苯、联苯、6-甲氧基-2-乙酰萘、戊基氰基三联苯、乙醛、甲醛、乙醇钠、、乙酸乙酯、乙醇(无水)、复盆...
如图,上面是氯乙醇,下面是2-氯乙醇 主要区别在于氯原子连接的碳原子不同,氯乙醇的氯原子连接的是一号碳原子,所以叫1-氯乙醇,但1通常省略,2-氯乙醇的氯原子连接的是2号碳原子,所以叫2-氯乙醇,为了...

 DuPont子公司在美国建造工业规模生物乙醇装置
DuPont子公司在美国建造工业规模生物乙醇装置
DuPont子公司在美国建造工业规模生物乙醇装置
Dupont子公司DupontDanisco纤维素乙醇(DDCE)公司将在美国爱荷华州Nevada建造一套工业规模生物炼油厂,用于由纤维素生产燃料级乙醇。纤维素将从玉米秸秆和麦草(包括干玉米棒、茎和收获后剩余的叶子)中得到。
 天冠“秸秆乙醇”“生物柴油”示范项目通过验收
天冠“秸秆乙醇”“生物柴油”示范项目通过验收
天冠“秸秆乙醇”“生物柴油”示范项目通过验收
由河南天冠集团承担的河南省重大科技专项“年产3000吨秸秆纤维乙醇关键技术研究及产业化示范”和河南省重大科技攻关计划“生物柴油清洁生产关键技术开发和产业化示范”项目,10日通过省科技厅专家组验收。
棉纤维作为含碳量较高的纤维素纤维,其回收利用前景广阔。回收的废旧棉纤维可以经碳化制备碳微球,作为吸附碳材料使用;也可以降解为葡萄糖制备生物乙醇,实现废旧棉纤维的资源化;力学性能较好的棉纤维还可二次纺丝,解决纺织原料紧缺的问题。然而棉纤维的聚合度和结晶度较高,大大限制了其高效回收利用。磷酸法低耗环保、节约资源,是棉纤维回收利用的有效途径。本文介绍磷酸在废旧棉纤维回收领域的应用。

磷酸法制备再生纤维素材料
磷酸/多聚磷酸液晶纺丝
P2O5质量分数达到一定范围的磷酸/多聚磷酸体系是纤维素的优良溶剂,在纤维素质量分数达到一定值后,可形成各向异性溶液,有着独特的液晶性。这种磷酸/多聚磷酸溶液体系在合适的温度和工艺条件下可以制备出高强、高模纤维。
磷酸活化炭纤维
传统的碳材料制备方法具有碳颗粒成长快速,微观结构易于控制,产品质量高、纯度大等特点。有研究表明,磷酸可促进纤维素脱水,加上磷酸的阻燃及交联作用降低了纤维的大量裂解和碎片的逸散,使纤维的碳残留率提高。
吸附性碳材料
采用磷酸法活化以棉秆为原料可以制取具有优良脱色性能的活性炭,且活性炭收率较高。
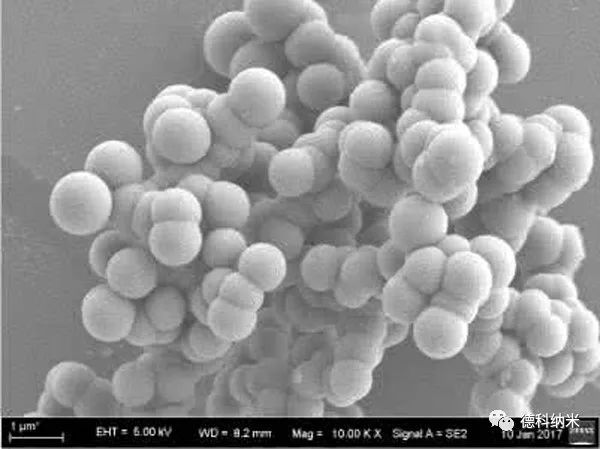
以纤维素为基底的复合材料
全纤维素复合材料
全纤维素复合材料是完全可再生材料,不仅解决了复合材料制备过程中界面相容性的问题,降低制备的能耗,还具有优异的机械性能。全纤维素复合材料的基体和增强相为同种材料,通过控制磷酸溶解条件可实现纤维素的表面部分选择性溶解,溶解部分成为增强相与未溶解部分进行复合,形成具有优异机械性能的材料。

纤维素与聚丙烯复合材料
聚丙烯/纤维素复合材料是一种健康环保的材料,可在许多场合替代天然木材和其他化学建材。棉纤维的纤维素含量高达95%,不含木质素、半纤维素等杂质。将棉纤维应用于智能化食品包装,是废旧棉纤维工业化的重要途径。将少量的纤维素混入商品塑料可以显著改善包装袋的性能。食品包装塑料的基础设计研究侧重于塑料基复合材料。

生产制备燃料乙醇
废旧棉织物中的棉纤维,纤维素含量高达50% ~95%,其作为纤维素纤维可代替可再生的生物质来源来提炼乙醇。浓磷酸预处理废弃棉织品所得再生纤维素进行酶水解,水解率可达92%以上。此外,浓磷酸预处理后的再生纤维素对酶的吸附能力显著提高,增加了酶的接触位点,可使纤维素水解率提高3.22倍。以浓磷酸预处理废弃棉织物,获得再生纤维素,对其进行酶水解,为废弃棉织物高效资源化利用提供有益的技术支撑。
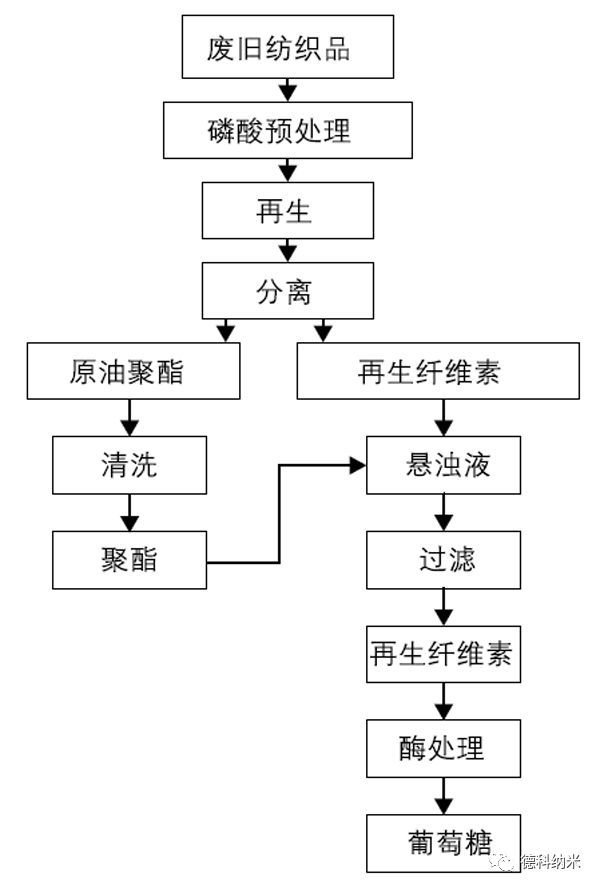
磷酸的回收
磷酸既能水解纤维素也能溶解纤维素。纤维素在磷酸中的溶解是可逆的:其溶解过程是磷酸和纤维素中羟基相互作用形成磷酸酯纤维素;其析出过程是水与磷酸根的结合能力强于纤维素与磷酸根的结合能力,故样品加水后纤维素因溶解能力降低而析出。磷酸在破坏棉纤维结构,降低废旧棉纤维结晶度和聚合度生成再生纤维素的同时,也可实现浓缩精制后重复利用。即磷酸与纤维素的反应是可逆的,在形成再生纤维素的同时可实现磷酸的回收。可以说,磷酸法低碳环保,能耗小,是颇具发展前景的回收废旧棉纤维的方法之一。(了解本文详细内容,请关注:张美玲,史晟,侯文生,等.磷酸在废旧棉纤维回收领域的应用[J].纺织导报,2017(07):63-66.)
文章来源于网络;如有涉及版权问题,请联系小编删除。
第1章 导论:生物乙醇的重要性与厌氧细菌及其纤维小体的贡献1
参考文献5
第2章 纤维小体传奇:早期研究7
2.1 引言7
2.2 发现过程8
2.3 后续研究11
2.4 证据14
2.5 未来方向21
2.6 结论22
参考文献22
第3章 粘连蛋白与对接蛋白的相互作用33
3.1 引言34
3.2 粘连蛋白与对接蛋白的系统发生多样性34
3.3 粘连蛋白模块34
3.4 对接蛋白模块37
3.5 粘连蛋白?对接蛋白复合物的结构37
3.6 结合的专一性39
3.7 粘连蛋白?对接蛋白相互作用及纤维小体复合体41
3.8 结论41
参考文献42
第4章 纤维小体的催化组分49
4.1 纤维小体:超分子复合体49
4.2 结构模式50
4.3 结论58
参考文献59
第5章 热纤梭菌降解纤维素:微生物角度67
5.1 引言67
5.2 热纤梭菌的生理生态学68
5.3 纤维素水解机理68
5.4 生物能学69
5.5 纤维素酶合成的调控70
5.6 黏附机制72
5.7 生物膜反应73
5.8 最新前沿75
参考文献77
第6章 热纤梭菌纤维小体的新组分及基因组序列分析82
6.1 引言82
6.2 热纤梭菌降解纤维素过程83
6.3 热纤梭菌基因组中的纤维小体基因88
6.4 基因组中潜在的纤维小体纤维素酶基因89
6.5 基因组中组成纤维小体的半纤维素酶基因90
6.6 基因组中的糖苷酶基因91
6.7 完整的果胶酶系统92
6.8 基因组中的其他水解酶与酯酶基因92
6.9 纤维小体中非催化组分的基因93
6.10 碳水化合物结合结构模块的功能93
6.11 纤维小体蛋白的模块组装?多功能酶93
6.12 在纤维素上培养时实际存在的纤维小体组分95
6.13 纤维素水解中未确定功能组分Cel8A的可能作用96
6.14 纤维小体组成随底物的变化96
6.15 非纤维小体纤维素酶系的增补作用96
6.16 哪些纤维糊精是纤维素水解的产物97
6.17 结论99
参考文献99
第7章 噬纤维梭菌纤维小体106
7.1 引言106
7.2 噬纤维梭菌纤维小体的组成和结构107
7.3 纤维小体基因的组装109
7.4 纤维小体异质性的亚基组成109
7.5 纤维小体基因的转录和表达调节110
7.6 纤维小体的设计110
7.7 利用小型纤维小体研究协同作用111
7.8 DNA重排方法改进纤维小体酶的特性111
7.9 纤维小体基因的异源表达112
7.10 CBD的应用112
7.11 结论与注释112
参考文献113
第8章 约氏梭菌与白色瘤胃球菌的纤维小体117
8.1 约氏梭菌的纤维小体117
8.2 白色瘤胃球菌的纤维素降解系统125
参考文献127
第9章 纤维小体嵌合体:研究纤维小体功能及其潜在生物技术应用的有效工具131
9.1 引言131
9.2 纤维小体嵌合体的概念133
9.3 利用纤维小体嵌合体来研究解纤维素梭菌产生的纤维小体135
9.4 未来前景139
9.5 未来生物技术的应用140
9.6 结论141
参考文献141
第10章 黄色瘤胃球菌纤维小体的组织结构144
10.1 引言144
10.2 黄色瘤胃球菌中纤维小体的组织架构145
10.3 黄色瘤胃球菌降解植物细胞壁的其他特征152
10.4 结论153
参考文献154
第11章 解纤维梭菌:纤维小体及其纤维素水解158
11.1 引言158
11.2 纤维小体的酶组分159
11.3 纤维小体脚手架蛋白164
11.4 纤维小体166
11.5 非纤维小体的蛋白质168
11.6 纤维素降解168
11.7 结论171
参考文献172
第12章 热纤梭菌纤维素酶生物合成调控179
12.1 碳源调控179
12.2 单个纤维素酶组分的调节180
12.3 生长速率的影响180
12.4 纤维素酶基因簇181
12.5 负调节182
12.6 结论183
参考文献183
第13章 厌氧真菌的纤维小体185
13.1 引言185
13.2 厌氧真菌纤维小体的酶组分190
13.3 非催化的对接蛋白结构域(NCDD)和脚手架蛋白197
13.4 厌氧真菌中纤维素降解酶的基因组结构199
13.5 结论199
参考文献200
索引209
-------- 1、 研究方向主要集中在3个方面:生物环境、生物能源、生物纳米(1)生物环境重点研究植物对重金属(Pb\Cd\Hg\Cu\Tb)元素吸附作用的机理及转基因技术,培育转基因抗逆植物、微生物新品种。(2)生物能源 重点研究木质纤维素制取生物乙醇燃料原理及工艺,培育转基因速生能源林、高产优质纤维素酶、半纤维素酶工程菌及高活性纤维素酶酵母工程菌和产朊假丝酵母工程菌新品种。主要包括3方面: A、 逆高产优质纤维素酶、半纤维素酶工程菌、高活性纤维素酶酵母工程菌和产朊假丝酵母工程菌等培养与研究。 B、并行糖化共发酵(SSCF)技术研究:糖化与C6、C5共发酵,消除纤维素酶受葡萄糖和纤维二糖的终产物抑制、半纤维素酶受木糖等五碳糖终产物抑制,提高纤维素酶、半纤维素酶的酶解效果,并降低酶制剂的用量。 C、细胞表层显示技术研究:在酵母细胞表层蛋白(α-凝聚素等)基因上植入纤维素酶等基因,提高醇化效率,降低成本,菌体和酵素持续高浓度反复利用。(3)生物纳米 重点对生物纳米材料及载体在植物体的组织、细胞、细胞器中的作用机理及相关应用技术研究(生物纳米转基因载体、生物纳米传感信息及生物纳米芯片);同时,开展生物纳米医药研究(药用蛋白质、药用酶、功能多肽及活性次生代谢物合成酶基因克隆及工程菌发酵技术)。
2、 科研课题 5年内争取科研课题经费3000万元,其中纵向课题1000万元、横向课题2000万元。纵向课题:国家“948”课题、国家“973”课题、国家“863” 课题、国家“自然科学基金”课题等。横向课题:美中合作课题、欧盟合作课题、中韩合作课题、高科技企业合作课题等。
3、 引进人才规划国内、国外引进优秀人才10人次,包括长期聘任、合作,短期聘任、合作等多种形式。拟聘的主要科学家: Ben Koopman 博士:美国 University of Florida (Gainesville, FL) 环境工程教授,美国国家科学基金会工程研究中心环境纳米分离技术组带头人,污水处理专家,尤其在污水生物处理技术及污水处理系统之监测--控制--自动化 (ICA) 等方面颇有造诣。 Changwon Kim 博士:韩国 Pusan National University (Pusan, South Korea) 环境工程教授,International Water Association (IWA) 监测--控制--自动化 (ICA)专业委员会主席,尤其在污水处理系统之动态监控和管理自动化方面颇有成就。 Lonnie O. Ingram 博士:美国佛罗里达大学微生物与细胞科学系教授、美国国家科学院院士,可再生化学品与燃料研究中心主任,是著名生物燃料和细胞工程专家, 获得过包括木质纤维素生产燃料酒精在内的有关生物质酶解与发酵微生物基因工程和细胞工程的十多项发明专利,多次受到联邦、州、和学校各级的嘉奖,特别是美国国会参、众两院以及总统的表彰。 Wayne H. Smith博士:美国佛罗里达大学森林系教授、林学院院长、生物能源研究中心主任, 国际知名的生物能源专家,长期从事生物能源(特别是木本能源)的研究开发与应用推广工作,并有丰富的国际合作与咨询经验,包括欧洲生物能源与环境论坛特约专家,联合国粮农组织印度生物能源教育项目顾问,泰国皇家科学院生物能源项目顾问,美国国际开发署生物能源援助项目顾问,美国能源部秘鲁生物质资源评估顾问等。2100433B