无性生殖
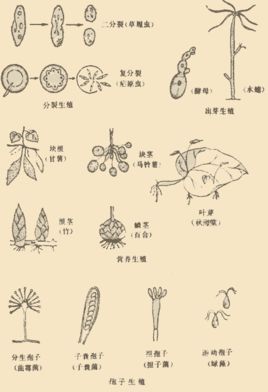
无性生殖是一类不经过两性生殖细胞的结合,由母体直接产生新个体的生殖方式。 可以分为分裂生殖(细菌及原生生物)、出芽生殖(酵母菌、水螅等)、孢子生殖(蕨类等)、营养生殖(草莓匍匐茎等),具有缩短植物生长周期,保留母本优良性状的作用。像水螅出芽,和无心插柳柳成荫等生命现象,其实就是生物在进行无性生殖。
-
选择特殊符号
选择搜索类型
请输入搜索
无性生殖是一类不经过两性生殖细胞的结合,由母体直接产生新个体的生殖方式。 可以分为分裂生殖(细菌及原生生物)、出芽生殖(酵母菌、水螅等)、孢子生殖(蕨类等)、营养生殖(草莓匍匐茎等),具有缩短植物生长周期,保留母本优良性状的作用。像水螅出芽,和无心插柳柳成荫等生命现象,其实就是生物在进行无性生殖。
椒草:椒草的叶片养在水里就能够形成新的根系,发出芽和根,并长成新的植株。
马铃薯:马铃薯块茎发芽,可移栽。

土豆、红薯、山药(用块茎繁殖) 杨树、柳树、玫瑰、月季(扦插) 苹果、梨、、核桃、等等优良品种的培育(嫁接) 会产生气生根的植物可以用压条法,如榕树、夹竹桃 植物组织培养也属于无性生殖,如许多农作物新...
蕨类是靠孢子无性繁殖,被子植物靠有性繁殖。蕨类植物是植物中主要的一类,是高等植物中比较低级的一门。是最原始的维管植物。大都为草本,少为木本。孢子体发达有根、茎、叶之分,不具花,以孢子繁殖。世代交替明显...
孔雀开屏是由体内神经和激素调控的行为,一方面是雄孔雀成熟了,绚丽的尾羽是激素调节的结果,生物学上称为"第二性征";另外一方面,开屏必须受到外界刺激才表现,如两只雄鸟相遇或者用鲜艳的花布刺激它,因此,这...

 紫薇优良无性系生长和光合特征分析
紫薇优良无性系生长和光合特征分析
紫薇优良无性系生长和光合特征分析
通过评价紫薇优良无性系的生长和光合特性,为其合理配置和科学栽植提供理论依据。以引进和选育的5个紫薇优良无性系为试验材料,进行相关特征测定,比较5个紫薇优良无性系的生长和光合特征差异。研究结果表明:5个无性系生长和光合生理特征差异达到了极显著水平(P<0.01),地径、叶长、叶宽、枝长、枝基径、叶绿素a、叶绿素b等最小值分别为2.41 cm、6.13 cm、3.75 cm、0.72 m、0.95 cm、0.95 mg/g和0.24 mg/g,其中‘速生紫薇’地径(4.86 cm)、叶长(9.99 cm)、叶宽(6.13 cm)、枝长(1.77 m)、枝基径(1.80 cm)、叶绿素a(1.68 mg/g)、叶绿素b(0.49 mg/g)等均较突出;5个无性系光饱和点(LSP)在1672.05μmol/(m2·s)以上,较喜光,光补偿点(LCP)在20~31μmol/(m2·s)之间,最大净光合速率(Pnmax)大小排序为‘鄂薇2号’[25.43μmol/(m2·s)]>‘速生紫薇’[19.67μmol/(m2·s)]>‘鄂薇4号’[18.38μmol/(m2·s)]>‘鄂薇1号’[16.64μmol/(m2·s)]>‘鄂薇5号’[11.17μmol/(m2·s)];相关分析表明叶片大、叶绿素含量高则耐荫性稍强,叶片暗呼吸速率对地径影响明显。在实际栽培中,可根据生长和光合特性,同时结合培育目标合理配置5个优良无性系。
紫薇部分速生无性系幼苗生长规律研究
紫薇部分速生无性系幼苗生长规律研究
为了选育出适合苗圃培育的紫薇无性系品种,以泰山林业科学研究院初选的10个速生性状比较明显的紫薇无性系扦插苗为材料,调查分析各无性系间幼苗苗高、50 cm地径的年生长量。结果表明,‘大华6号’苗高平均年生长量最大(124.3714 cm),地径平均年生长量最大(1.0514 cm);‘岱2号’苗高和地径平均年生长量次之,这2个无性系是具有明显速生性状的紫薇无性系品种。
海绵动物的生殖为无性生殖与有性生殖两种方式。无性生殖是以出芽生殖为主,多发生在海产种类中。出芽时亲本的变形细胞,特别是一些原细胞由中胶层迁移到母体的顶端表面聚集成团,然后发育成小的芽体,随后脱落到底部发育成新海绵,或与母体相连形成群体。淡水海绵及少数海产种类在一定条件下可以形成芽球(gemmule),也被认为是一种无性生殖,个体中的原细胞摄食了大量的物质之后聚集成团,外面包围一层造骨细胞。在原细胞团之外自行分泌一层保护膜,其成分类似于海绵丝,以保护内部的芽球细胞,之后造骨细胞分泌一层双盘状或针状的骨针,使芽球具有很强的抵抗恶劣环境的能力。一个海绵动物可以形成许多的芽球。以后当外界条件适当时,芽球内的细胞通过微孔(micropyle)释放出来,再形成一个新个体。海产种类的芽球外面包有海绵丝,具有或不具有骨针。少数种类的芽球不具有海绵丝。
海绵动物的再生也被认为是一种无性生殖,许多种类的海绵都有很强的再生能力,例如白枝海绵,它的身体碎片只要大于0.4mm,并带有一些领细胞,就能再生成一个新个体,这是由于海绵动物的细胞具有较强的聚合能力与识别能力。也有人将海绵动物的身体用机械方法压碎,将细胞分离,再用纱布过滤,其滤液中的分散细胞再放入海水中培养,结果分离的细胞又重新聚合,并分别迁移到正确的位置上,最后形成一个或几个新的个体。还有一个经典的实验是由Galtsoff(1925)所进行的,他用两种不同属的海绵做实验,即一种是细芽海绵(Microciona),其细胞具红色素;另一种是Haliclona,其细胞内具黄色素,他将两种预先分离成悬液的海绵细胞混合在一起,起初两种细胞随机聚合,但很快两种细胞按种彼此分开,分别形成红色细胞群及黄色细胞群,以后两种不同的细胞群各自分化,最后形成细芽海绵(红色素细胞)和Haliclona(黄色素细胞)两种新个体。以后也发现许多淡水海绵及海产的海绵都有此特性。后来有人用实验证实了海绵细胞表面的一种大分子量的糖蛋白是海绵细胞的识别分子,它具有种的特异性,所以同种的细胞能聚合,不同种的细胞相分离。同种细胞的聚合能力使它能再生及组成新的个体。
除了四射海绵(Tetractinellida)之外,海绵动物均能行有性生殖。大多数种类为雌雄同体(hermaphordite),但精子与卵常不在同一时期成熟。少数种类为雌雄异体(dioecism)。生殖细胞由中胶层中的原细胞形成,有时领细胞也可以失去鞭毛及原生质领而变成精原细胞(spermatogonia),再分裂形成精子。精子成熟后随水流排出体外,并随水流进入其他个体的鞭毛室。有人观察到某些热带地区的海绵能突发性的释放精子于海水中,形成一条乳白色的云雾状的精子带,其长度可达2—3m。一个海绵释放了精子常可诱导周围海绵也释放精子。释放出的精子随水流进入其他个体的鞭毛室之后,再进入领细胞。这时领细胞失去领及鞭毛,携带着精子到中胶层与卵融合而成受精卵。大多数海绵动物的受精卵是在体内发育。一些海绵动物的胚胎发育由于胚层的逆转而有很大的特殊性,不同纲的海绵胚胎发育过程也不尽相同,这主要表现在海绵动物可以形成两种不同类型的幼虫。在钙质海绵中,形成中空的两囊幼虫(amphiblastula),在寻常海绵纲中形成实心的实胚幼虫(parenchymula)。
钙质海绵,例如白枝海绵或毛壶的受精卵在母体的中胶层中发育,当受精卵经细胞分裂形成16个细胞时,构成动物极的为8个小细胞,构成植物极的为8个大细胞。动物极的小细胞分裂较迅速,分裂到一定数目的细胞之后,形成了一个具有囊胚腔的囊胚,小细胞面向囊胚腔的一端都长出一根鞭毛,以后小细胞经大细胞间的开口向外翻出,结果小细胞的鞭毛移到表面,形成了一个一端有鞭毛、一端无鞭毛的两囊幼虫。两囊幼虫随水流离开母体,在水中游泳一段时间之后经过小细胞的内陷、或大细胞的外包,或两种方法的联合而形成了两层细胞的原肠胚状,并固着在底部。原来动物极的具鞭毛的小细胞形成了成体的胃层(领细胞层),原来植物极的大细胞形成了成体的皮层(扁平细胞层),再由两层细胞共同形成中胶层及变形细胞。海绵动物的这种细胞分化与分层,与所有其他多细胞动物的胚层分化不同。在除海绵动物之外的其他后生动物中,动物极的小细胞发育成成体的外层(即外胚层),植物极大细胞发育成内层(即内胚层)。钙质海绵动物的这种动物极与植物极细胞相反的分化现象称为逆转现象(inversion),因此它的两层细胞不称为外胚层与内胚层,而分别称为皮层与胃层。发育中的逆转现象是将海绵动物列为侧生动物的原因之一。
钙质海绵中特别是复沟型海绵在发育中也经过一个原始阶段的重演,即发育中经过一个单沟型的原海绵(Olynthus)阶段,再经过一个双沟型的发育阶段之后,最后才形成一复沟型的成体。 大多数六放海绵及寻常海绵发育中经过一个实胚幼虫,实胚幼虫的外表面除后端外均为具鞭毛的小细胞,以后具鞭毛的外层细胞移入内部,形成胃层,内部的变形细胞移到外面形成皮层。寻常海绵纲中,许多复沟型种类,发育时直接来自一个复沟型幼虫(rhagon),这种幼虫具有宽阔的基部、狭小的顶端、很大的海绵腔、很小的鞭毛室,经过体壁褶叠后发育成复沟型成体。
扦插属于无性生殖。依选取植物不同的营养器官作插穗,按取用器官的不同可分为枝插、根插和叶插三类。

外植体→脱分化( 适当植物激素的比例)(无光照)→愈伤组织→再分化(由愈伤组织成为幼苗或胚状结构)→营养生长,生殖生长(发育成完整的植物体)
植物细胞脱分化再分化的关键因素:植物激素用量的比例、激素的使用顺序、植物激素的浓度
繁殖方式:无性生殖