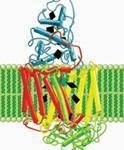
分子马达
分子马达(molecular motor),是美国康奈尔大学研究人员在活细胞内的能源机制启发下,制造出的一种马达。这种微型马达以三磷酸腺苷酶为基础,依靠为细胞内化学反应提供能量的高能分子三磷酸腺苷(ATP)为能源。
-
选择特殊符号
选择搜索类型
请输入搜索
分子马达(molecular motor),是美国康奈尔大学研究人员在活细胞内的能源机制启发下,制造出的一种马达。这种微型马达以三磷酸腺苷酶为基础,依靠为细胞内化学反应提供能量的高能分子三磷酸腺苷(ATP)为能源。
解螺旋酶:它利用分子机器,沿着DNA和 RNA运动.
肌动蛋白和动力蛋白:能够沿着轨道运动,轨道由同一亚单位(如肌纤蛋白和微管)重复而组成,是一种蛋白丝。
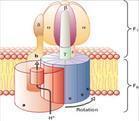
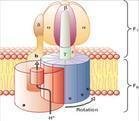
鞭毛:例如大肠杆菌之类的细菌,它们的运动依靠鞭毛,鞭毛扮演着螺旋桨的角色,在细菌细胞膜中做马达旋转,这个旋转的马达被一个跨膜的蛋白质浓度梯度所驱动,代替被ATP水解所驱动。
肌球蛋白:肌肉收缩。
F0F1三磷酸腺苷合成酶(ATPase): 位于细胞膜或线粒体膜上。利用电化学质子的梯度合成三磷酸腺苷(ATP)。
分子马达靠很小的增值来工作,转入蛋白质构象的改变从而能进行有引导的运动,它需要一条引导马达装配的运动轨道进行有规则的运动来通过一段距离。实际上,我们以前已经遇到过一类分子马达,它利用我们即将涉及的分子机器,即沿着DNA和RNA轨迹运动的解螺旋酶。沿重复同一亚单位组成的蛋白丝(如肌纤蛋白和微管)--在高度亲缘关系和低亲缘关系间的马达蛋白质的循环,是为使丝状轨道响应于ATP的结合,水解。ATP的每一次结合、推动、释放,都是产生运动的机制。
也存在一种完全不同的策略,就象大肠杆菌之类的细菌那样用来产生运动,一套鞭毛扮演着螺旋桨,在细菌细胞膜中做马达旋转,这个旋转的马达被一个跨膜的蛋白质浓度梯度所驱动,代替被ATP水解所驱动,一套蛋白质浓度梯度去转动运动的机理类似于ATP合成酶的F0亚基的作用.但是,储存生化能量的主要模式都是ATP和离子浓度梯度,被渐进式的利用去驱动有机分子运动.
真核细胞含有三种主要的马达蛋白家族:肌球蛋白、kinesins蛋白和动力蛋白。初一看,这些蛋白家族好像彼此很不同。在肌肉中的肌球蛋白,开始时被描绘成有它自己的作用基础,沿着肌纤蛋白的丝运动,肌肉肌球蛋白包括两个拷贝,它们都有一个87kd分子团的重链,一个必需的轻链,和一个起调节作用的轻链。人类基因似乎能编码超过40种截然不同的肌球蛋白,在肌肉收缩中有些功能和另一些参与不同种类的其他过程。kinesins蛋白在蛋白质、囊泡和沿微管的细胞器转运中起作用,包括染色体分离。kinesins蛋白常包括两个拷贝,一个是重链,一个是轻链,它的重链大约只有肌球蛋白长度的一半。人类基因至少能编码40种kinesins蛋白。在一些真核细胞中,动力蛋白能驱动纤毛和鞭毛的运动,和其他作用蛋白相比,动力蛋白较大,有个大于500kd的分子团重链,人类基因似乎能编码大约10种动力蛋白。
比较肌球蛋白,kinesins和动力蛋白的氨基酸序列,并没有出现这些蛋白质家族之间有意义的关系,但是,在确定它们的三维结构之后,肌球蛋白和kinesins家族的成员之间被发现有显著的相似性,特别是肌球蛋白和kinesins都包含同源的P-环NTP酶核心部位,这些在G蛋白中也存在。动力蛋白重链的序列分析揭示出它是P-环NTP酶的AAA子家族的一个成员,我们以前在19S蛋白解体系统中遇到过该子家族,动力蛋白有6个序列编码一个沿着它的长段排列的P-环NTP酶整环。从而,可以利用有关G蛋白和其他P-环NTP酶的知识来分析这些马达蛋白的运动机理。
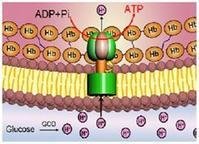
分子马达 (molecular motor)是美国康奈尔大学研究人员在活细胞内的能源机制启发下,制造出的一种马达。这种微型马达以三磷酸腺苷酶为基础,依靠为细胞内化学反应提供能量的高能分子三磷酸腺苷(ATP)为能源。
研究人员把金属镍制成的螺旋桨嫁接到三磷酸腺苷酶分子中轴上。当它们被浸于ATP溶液后,其中5个分子马达转动了起来,转速达到每秒钟8转。据介绍,这种马达只有在显微镜下才能被观察到,其镍螺旋桨长750纳米(一纳米为十亿分之一米)。根据拍摄到的画面,研究人员可以看到一个尘埃粒子先被旋转的螺旋桨吸入、再被甩出的情景。
分子马达,又名分子发动机,是分布于细胞内部或细胞表面的一类蛋白质,它们的构象会随着与ATP和ADP的交替结合而改变,ATP水解的能量转化为机械能,引起马达形变,或者是它和与其结合的分子产生移动。就是说,分子马达本质上是一类ATP酶。例如肌肉中的肌球蛋白会拉动粗肌丝向中板移动,引起肌肉收缩。而另外两种分子马达:驱动蛋白和动力蛋白,它们能够承载着分子“货物”-------------如:质膜微粒,甚至是线粒体和溶酶体,在由微管构成的轨道上滑行,起到运输的作用。
一类由生物分子组成,具有马达(发动机)功能的分子机器,能够象马达一样依赖于微管推进细胞器运动,故称为分子马达。常见的分子马达多为蛋白质,其家族有:驱动蛋白、动力蛋白和依赖于微丝的肌球蛋白,利用ATP水解得到的能量(化学能)推动马达转动(动能),从而推动细胞器运动。
1. 极数反映出电动机的同步转速,2极同步转速是3000r/min,4极同步转速是1500r/min,6极同步转速是1000r/min,8极同步转速是750r/min。 绕组的一来一去才能组成回路,也...
台湾东力要比城邦成立的早,已经有40年的历史了,台湾东力一直致力于减速马达的研发,所有配件包括润滑油都是国外进口的,马达的品质要比城邦好得多。搜索台湾东力电机这个关键词会在页面上看到“台湾东力电机股份...
你需要两块磁铁,铜线,连块细薄铁板,一块铁板对折,下端劈叉,再将三个支上均匀缠上,另一块作外壳,上附磁铁,即可

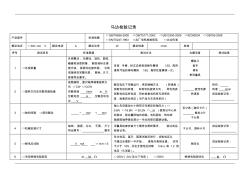 马达检验记录表
马达检验记录表
马达检验记录表
马达检验记录 产品型号 标准依据 □GB/T6656-2008 □GB/T5171-2002 □GB12350-2009 □IEC60034 □GB755-2008 □SN/T0247-1993 □出厂电机检验规范 □企业标准 额定电压 □DC□AC V 额定电流 A 额定功率 W 额定转速 r/min 其他 序号 测试项目 标准限值 测试方法 仪器设备 测试结果 1 □外观质量 外观整洁,无锈蚀、划伤、裂纹、 磕碰和涂层剥落, 颜色和标志清 楚无误,紧固件连接牢固, 引线 或接线柱完整无损, 颜色、尺寸、 规格符合要求。 目测 手感,标志应承受湿棉布擦抹 15S,再用 浸有汽油的棉布擦抹 15S,每秒往复擦抹一次。 螺丝刀 扳手 棉布 常用量具 2 □旋转方向及空载性能检查 按图接线,面对轴伸端看旋转方 向 □CW □CCW 空载
 鼓风机马达
鼓风机马达
鼓风机马达
你知道鼓风机马达吗 鼓风机简介 鼓风机主要由下列六部分组成: 电机、空气过滤器、鼓风机本体、 空气室、底座(兼油箱)、滴油嘴。鼓风机靠汽缸内偏置的转子偏心 运转,并使转子槽中的叶片之间的容积变化将空气吸入、 压缩、吐出。 在运转中利用鼓风机的压力差自动将润滑送到滴油嘴, 滴入汽缸内以 减少摩擦及噪声, 同时可保持汽缸内气体不回流, 此类鼓风机又称为 滑片式鼓风机。 鼓风机历史 扇、吹管和皮囊,最早用于强制鼓风的器具是扇和吹管。古埃及 金匠曾使用带陶风嘴的吹管,印加人有时用 8~12根铜管同时吹炼。 稍后 ,发明了用兽皮制作的鼓风皮囊,囊的两端分设风管和由操作者 手控的进风口。 这种简陋的鼓风器在近代仍在一些地区使用。 埃及第 十八王朝勒克米尔( Rekhmir,约公元前 1450年)墓的壁画中已绘有 罐状脚踏鼓风器的图象。 进风时,操作者用绳索拽起皮囊, 随后踩下, 将风鼓入炉内,每炉配备
从测量的角度看,实验科学的发展其实就是一个不断提高测量精度的过程。精度提高一步,科学就前进一步。这一点在分子生物物理中也不例外。有一类生物分子,一般称为分子马达,利用 ATP 水解产生的能量做轨道运动,完成其重要功能。
以 DNA 解旋酶为例,一般的理解是:解旋酶消耗一个 ATP,打开一对碱基,并沿着 DNA 向前移动一步。要定量刻画解旋酶的分子机理,测量精度至少要达到 0.3 nm,这是 DNA 双螺旋中碱基对之间的最小距离。单分子荧光共振能量转移(FRET)是研究解旋酶分子机理的重要工具,它应用偶极子之间相互作用对距离敏感的原理,测量分子马达的运动。理论上讲,FRET 可以测量 0.1 nm 的距离变化。然而,当将 FRET 应用于 DNA 双链的解旋研究时,很少能在室温下的水溶液中逼近这一极限,以至于很难区分本来应该有很大差别的解旋酶(图1a)。
中国科学院物理研究所/北京凝聚态物理国家实验室(筹)软物质与生物物理实验室李明研究组多年来致力于发展单分子技术研究生物大分子的动力学,在DNA凝聚和染色质组装(JACS 2006,PRL 2012,JACS 2013,Mol. Cell 2016)、分子马达(EMBO J. 2009,NAR 2014,NAR 2015)以及膜蛋白动力学(Nature Comm. 2016)等诸多方面取得了系列成果。这些研究的主线是通过高精度的单分子测量获取生物大分子的精细动力学信息。最近,该研究组的陆颖副研究员与博士生林文霞、马建兵等又将单分子荧光的测量精度提高了一大步。他们根据弹性力学计算,设计了一种称为“纳米张力器(nanotensioner)”的 DNA 结构,将一小段 DNA 弯成一张弓,利用 DNA双 螺旋结构的弯曲弹性,将张力直接作用于待解开的 DNA 双链岔口,将 DNA 单链撑开并抑制其热涨落(图1b)。
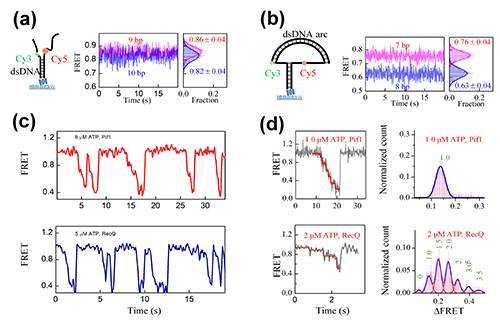
图1. (a)常规FRET实验中用带自由单链的DNA分岔结构作为底物,两个荧光分子的间距增加一个碱基对时,FRET效率只改变0.04。(b)用DNA张力器将自由单链撑开,两个荧光分子的距离增加一个碱基对时,FRET效率改变0.13。(c)常规FRET实验中,Pif1与RecQ的解旋数据类似,无法区分。(d)高精度实验中,两个解旋酶表现出完全不同的动力学性质。
他们将该方法应用于研究人源Pif1和大肠杆菌RecQ解旋酶。以往由于精度有限而看起来相似的解旋信号在新实验中表现得迥然不同。当然,表观上的不同并不意味着这些解旋酶完全不相干。高精度下测得的不同点,实际上隐含着更深层次的共同点。基于这个思路,他们提出了一个带普适意味的分子模型:解旋酶一次消耗一个ATP,破坏一对碱基的氢键;Pif1的氢键断裂与碱基释放同步,看起来一次解开一对碱基;RecQ的氢键断裂与碱基释放不同步,看起来一次解开随机个数的碱基对。该普适模型可以更准确更细致地刻画多种解旋酶的结构与功能的关系(图2),加深了我们对分子马达的理解。这一研究成果发表在最新一期Physical Review Letters(PRL 119, 138102 (2017))上。

图2. (a)基于高精度实验结果提出的解旋酶的普适解旋模型:解旋酶消耗一个ATP,破坏一对碱基的氢键,但并不立即释放打开的碱基。不同的解旋酶释放碱基的速率依赖于其自身的结构,产生不同的表观解旋动力学特征。(b)模特卡罗模拟表明,通过调节碱基释放的速率,该普适模型可以解释Pif1和RecQ不同的解旋步进(左列)及其在每一步的等待时间(右列)的统计分布规律。
该工作得到了国家自然科学基金委(11674382, 11574382, 11574381),中国科学院前沿科学重点项目(QYZDJ-SSW-SYS014)以及中国科学院青年创新促进会的大力支持。
编辑:Cloudiiink